Introducción
Las enfermedades parasitarias son un problema global que limita la salud veterinaria y el rendimiento productivo del ganado. Las garrapatas son el grupo más importante de patógenos que causan enfermedades a los bovinos. En específico Rhipicephalus microplus (R. microplus), es un ectoparásito hematófago obligado, el cual, es considerado como uno de los principales factores que limitan la ganadería en los trópicos y subtrópicos a causa de los daños directos e indirectos que ocasiona, dentro de los cuales, destaca su capacidad vectorial y expoliativa de transmisión de agentes infecciosos como Babesia bigemina, (B. bigemina), Babesia bovis (B. bovis) y Anaplasma marginale (A. marginale) (Solorio et al., 1999; Rodríguez-Vivas, 2004; de la Fuente et al., 2007). En relación, los ixodicidas son las herramientas más utilizadas para el control de las garrapatas (George et al., 2004). Sin embargo, el uso inadecuado se ha convertido en la principal presión de selección para estos organismos, que ha resultado en el surgimiento de poblaciones resistentes a estos compuestos químicos, aunado a la alta plasticidad del genoma de las garrapatas (Rosario-Cruz et al., 2009). Lo que ha puesto en relieve el desarrollo de diversos enfoques en la investigación, dirigidos hacia la búsqueda de nuevos blancos moleculares de la garrapata, inmunológicamente importantes para el desarrollo integrado de vacunas. En este sentido, muchos grupos de investigación proponen el control de vectores, basado en organismos acaropatógenos, como las bacterias Staphylococcus saprophyticus (S. saprophyticus) que son las causantes de la enfermedad del pico amarillo en las garrapatas de R. microplus (Miranda-Miranda et al., 2010). Esta infección, produce un exudado amarillento en las hembras del artrópodo, que bloquea el poro genital impidiendo la oviposición resultando en la muerte de la garrapata (Miranda-Miranda et al., 2010).
Por otra parte, el sistema inmunológico de las garrapatas tiene una amplia variedad de mecanismos de protección. De manera análoga que en insectos y otros artrópodos, la hemolinfa de las garrapatas es el fluido circulatorio homólogo a la sangre, donde se lleva a cabo el inicio de la respuesta inmunológica protectora. (Kauffman, 2007). Este sistema básicamente se divide en dos tipos de respuestas, una respuesta de tipo celular, que se lleva a cabo por un conjunto de células en movimiento denominadas hemocitos, que representan una de las primeras líneas de defensa. Una vez activados, los hemocitos responden mediante diversos mecanismos como fagocitosis, nodulación y encapsulación, y la producción de especies reactivas de oxígeno (ROS) (Strand, 2008). No obstante, existen varios patógenos que logran evadir los mecanismos del sistema inmune y colonizan los órganos del cuerpo de la garrapata. A la fecha, se sabe que el sistema inmune de los artrópodos posee una gran plasticidad que se refleja en el tipo de respuesta que genera. Esta respuesta es patógeno dependiente, e incluso presenta grandes diferencias entre bacterias Gram positivas y Gram negativas (Soneshine et al., 2013). Por otra parte, la respuesta inmune humoral consiste en moléculas efectoras solubles que incluyen defensinas, péptidos ricos en histidinas, factores de coagulación, lectinas, alfa-macroglobulinas (como las glicoproteínas), proteasas e inhibidores de proteasas, así como otras moléculas de reconocimiento, a los cuales, se suma un vasto repertorio de péptidos antimicrobianos (AMP’s), que son importantes en la respuesta inmune innata de diversos animales, insectos y plantas (Sonenshine et al., 2008; Iwanaga et al., 2005; Mcphee et al., 2005; Taylor et al., 2006). Sin embargo, en el vector R. microplus, la respuesta inmune de los AMP’s ha sido poco reportada.
Los AMP’s son polipéptidos genéticamente codificados y sintetizados en los ribosomas que forman parte de la respuesta inmune innata de los artrópodos (Wang et al., 2012). En la garrapata, estos AMP se sintetizan en el cuerpo graso y se liberan en la hemolinfa de manera rápida y efectiva como resultado a la invasión por microorganismos, mediante el reconocimiento de componentes superficiales de los microorganismos como el LPS y PGN presentes en las bacterias Gram positivas y Gram negativas. Estructuralmente los AMP’s son secuencias cortas (30-60 aminoácidos) y en su gran mayoría poseen carga positiva (Li et al., 2012). Las defensinas, son los tipos de AMP’s más estudiados en invertebrados y muestran un patrón clásico conservado de residuos de cisteína Cys-1-Cys-4, Cys-2-Cys-5, y Cys-3-Cys-6, los cuales, están unidos entre sí por tres puentes disulfuro que estabilizan la molécula y mantienen la estructura terciaria (Ganz, 2003). Asimismo, el mecanismo de acción de estos AMP’s esta embebido en la formación y organización de poros en la membrana del patógeno, mediante la inserción de los péptidos a través de interacciones electrostáticas, que resulta en la lisis por choque osmótico (Boman, 1995). Por lo anterior el objetivo principal de este trabajo, fue caracterizar la expresión de un nuevo péptido antimicrobiano (hebraina) y determinar su papel en la interacción patógeno-vector, durante la infección en la garrapata de ganado con S. saprophyticus y A. marginale, con el fin de proponer nuevos blancos terapéuticos moleculares e inmunológicos, que resulten en el desarrollo de nuevos fármacos y/o vacunas.
1. Materiales y métodos
1.1. Infección de garrapatas R. microplus con S. saprophyticus y A. marginale
La expansión de la colonia de garrapatas, se llevó a cabo a través de una infestación de un bovino macho con 1g de huevos equivalente a 20,000 larvas de R. microplus. Para la obtención de garrapatas infectas con anaplasma, se repitió el mismo procedimiento, pero en un bovino macho infectado con A. marginale, proporcionado por la Unidad de Anaplasmosis del CENID-SAI, del INIFAP. La comprobación de la infección con anaplasma, se realizó por amplificación de la secuencia del gen msp5 de A. marginale, a partir de DNAg extraído de carca de garrapatas infectadas, mediante ensayos de PCR punto final. Las infecciones de garrapatas con S. saprophyticus, se llevaron a cabo mediante la infestación de un bovino hembra como se describió anteriormente. Adicionalmente, se realizó un refuerzo con 0.5 g de larvas. A los 21 días post-infestación, se llevó a cabo la recolección de garrapatas ingurgitadas, y se lavaron 5x con una mezcla de antibiótico/antimicótico. Las infecciones con S. saprophyticus, se llevaron a cabo por baño en medio liquido selectivo en presencia de la bacteria. La confirmación de la infección, se realizó por la amplificación del gen 18s Ribosomal de S. saprophyticus con un PCR punto final. Los controles internos para todos los experimentos, fueron garrapatas ingurgitadas sin infección y tratadas bajo las mismas condiciones.
1.2. Determinación de la secuencia de la hebraina mediante análisis in silico
Al momento, la secuencia de la hebraina no se encuentra reportada en las bases de datos del genoma de R. microplus. Para la búsqueda y obtención de la secuencia, se realizaron análisis de BLASTn por homología con otras secuencias reportadas, en 3 diferentes bases de datos en los servidores (Pubmed, SwissProt y KEGG).
1.3. Diseño de oligonucleótidos
A partir de las secuencias obtenidas se diseñaron los oligonucleótidos en el programa PRIMER-BLAST de la base de datos del NCBI, tomando en cuenta los siguientes parámetros: obtención de fragmentos de 18 a 20 pares de bases, sin formación de dímeros, % de G-C no mayor al 55%, y temperatura de alineamiento no mayor a 2ºC. Con las secuencias de los oligos obtenidos, se realizó un BLASTn en la base de datos del NCBI, para la confirmación de identidad contra la secuencia original (Tabla 1).
Tabla 1
Oligonucleótidos diseñados a partir de la secuencia de ARNm de las secuencias obtenidas en la base de datos del NCBI. Se muestra la secuencia completa, el tamaño en pares de bases, la temperatura de alineamiento (TM), y el porcentaje de la composición de bases (% GC)
| Secuencia | Longitud | Tm | GC % | |
|---|---|---|---|---|
| Oligonucleótido Forward | AGTGTCTGCTTGGTTCTGGT | 20 | 59.16 | 57.89 |
| Oligonucleótido Reverse | AACTCCGTGTATGCAGCGAT | 20 | 59.82 | 52.38 |
1.4. Determinación de la presencia y expresión de la hebraina
Todas las extracciones de ácidos nucleicos, se realizaron a partir de hemolinfa de garrapatas infectadas con bacterias patógenas (A. marginale y S. saprophyticus), y de los controles sin infección. La extracción de DNA se realizó mediante la técnica de fenol: cloroformo: alcohol isoamílico, y el RNA con la técnica convencional de Trizol (Ambion Inc.), siguiendo las recomendaciones de fabricante. Para la determinación de la presencia de la secuencia de la hebraina en el genoma de la garrapata, se realizó un ensayo de PCR punto final a partir de DNAg extraído de células embrionarias de R. microplus. La determinación de los niveles de expresión relativa de hebraina en garrapatas infectadas e intactas, se realizó mediante el ensayo de RT-PCR con un kit One Step III (Invitrogen™|Thermo Fisher Scientific), siguiendo las indicaciones del fabricante. Las normalizaciones de la expresión relativa del mensajero de la hebraina, se realizaron por evaluación de la expresión del gen constitutivo de la Fosfolipido-hidroperoxidasa glutatión peroxidasa (phgpx), previamente reportado por Cossío-Bayúgar et al., en 2009. Todos los análisis densitométricos se realizaron con el programa Image J (National Institutes of Health).
1.5. Análisis estadístico
Los análisis estadísticos de realizaron utilizando los programas estadísticos SPSS versión 20.0 y Statistica versión 10.0. Las pruebas para la identificación de las diferencias entre los niveles de expresión entre los grupos experimentales se basaron en análisis de varianzas de dos factores (ANOVA de 2 vías), y pruebas de Tukey para identificar las diferencias significativas en la densitometría, considerándose los valores de P< 0.05 como estadísticamente significativos. Los datos obtenidos para los análisis estadísticos, son el resultado de tres pruebas distintas realizadas, cada una con triplicados.
1.6. Modelaje molecular
A partir de la secuencia obtenida, se realizaron modelajes en el programa SwissModel para la predicción de la estructura tridimensional de la hebraina, junto con la validación estereoquímica y energética. La comparación de la estructura y composición del AMP, basada en la estructura descrita para los péptidos antimicrobianos de insectos (Ganz, 2003), se realizó mediante un análisis de alineamiento con el programa MUSCLE, con las secuencias de otras defensinas reportadas. Estos alineamientos se realizaron tomado como referencia la defensina de Drosophila melanogaster, la cual, presenta regiones conservadas de residuos de cisteína como las clásicas reportadas para estos péptidos (Ganz, 2003).
1.7. Análisis filogenético
Para la determinación de las probables distancias evolutivas de las secuencias con alto porcentaje de identidad con la hebraina de R. microplus, se realizó un análisis filogenético. Estos análisis incluyeron una secuencia de un taxa ortólogo (grupos externos), para darle mayor validez. Todas las secuencias obtenidas, fueron alineadas mediante el programa Muscle del servidor ExPASy. Posteriormente, se llevó a cabo el análisis filogenético en el programa MEGA 7, generando una matriz de distancias, la cual, nos reporta el mejor árbol sugerido para el análisis de secuencias de los AMP’s. Finalmente, se construyó un árbol filogenético con 500 Bootstraps, y un grupo externo, por el método de Máxima Verosimilitud y el modelo evolutivo de Jones-Taylor-Thornton (Wollenberg 2000).
2. Resultados
2.1. Determinación de la presencia del gen de la hebraina en el genoma de R. microplus
La determinación de la presencia del gen de la hebraina en el genoma de R. microplus se llevó a cabo mediante ensayos de PCR punto final. Los resultados muestran un amplicon de 258 pb producto de tres diferentes extracciones de DNAg obtenido de células embrionarias de R. microplus mantenidas en medio de cultivo (Fig. 1).
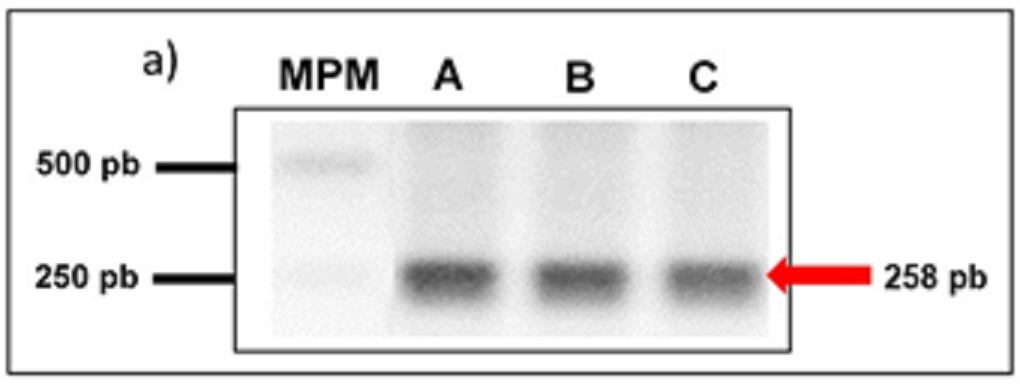 |
Determinación de la presencia del gen de la hebraina en el genoma de R. microplus mediante ensayos de PCR. Se observan tres productos de aproximadamente 258 pb, resultado de tres diferentes extracciones de DNAg obtenidos de tres cultivos de células embrionarias de R. microplus (carriles A, B y C). MPM: marcador de peso molecular 1000 pb. |
2.2. Determinación de la expresión relativa del gen de la hebraina
La determinación de la expresión relativa del gen de la hebraina se llevó a cabo mediante ensayos de RT-PCR. Los resultados muestran dos amplicones de aproximadamente 200 pb presentes en hemolinfa de garrapatas sin infección y hemolinfa de garrapatas infectadas con A. marginale (Fig. 2 panel a). Interesantemente, la hemolinfa de las garrapatas que fueron infectas con S. saprophyticus, no muestran expresión del mensajero (Fig. 2 panel a). La normalización de los amplicones obtenidos para el cálculo de la expresión relativa, se realizaron con gen phgpx. Los análisis densitométricos mostraron una ligera diferencia de expresión relativa entre los productos de las hemolinfas con infección de A. marginale de 71.46%, en comparación con los controles intactos, los cuales, presentan una expresión de 67.12% (Fig. 2 panel b).
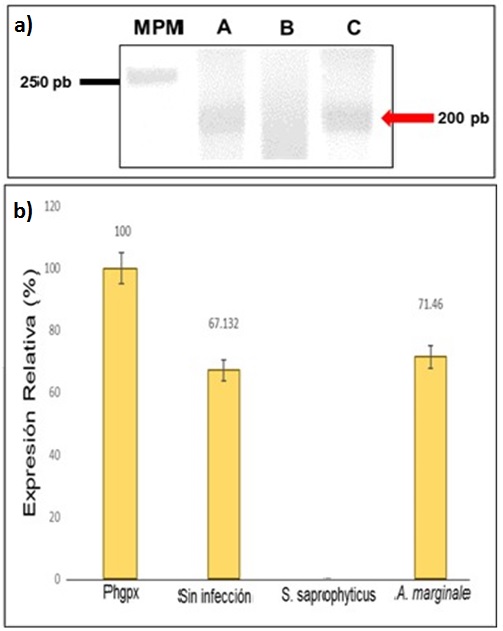 |
Determinación de la expresión relativa de hebraina. Panel a) Expresión relativa de la hebraina por RT-PCR. Carril A: indica los controles que corresponde a hemolinfa de garrapatas sin infección; Carril B: muestra la hemolinfa de garrapatas infectadas con S. saprophyticus; Carril C: indica la hemolinfa de garrapatas infectadas con A. marginale; MPM: marcador de peso molecular; Panel b) análisis densitométrico de la expresión relativa de la hebraina con el programa Image J. La gráfica muestra una expresión relativa de 67.132% en hemolinfa de garrapatas sin infección; de cero en hemolinfa de garrapatas infectadas con S. saprophyticus; y de 71.46% en hemolinfa de garrapatas infectadas con A. marginale. El phgpx representa el control constitutivo. Los análisis estadísticos (ANOVA) mostraron una P< 0.05 (*). |
2.3. Análisis estructural de la hebraina
La validación estereoquímica y energética de la estructura 3D, proporcionados por el programa Swiss-Model, mostraron valores dentro los parámetros establecidos para la calidad del modelo. Los resultados de la estimación del modelo global, mostraron un valor de 0.8 de un rango de 0 a 1, lo cual, indica un alto índice de confiabilidad del modelo, con un 75% de cobertura y 44.7% de identidad. En cuanto a la estimación de la calidad absoluta (QMEAN), basados en el Z-score, que estima el grado de natividad de las características del modelo, posee un valor de -0.93, valor que se encuentra dentro del rango establecido por el programa para los modelos de alta calidad.
Los análisis estructurales muestran diferentes regiones altamente conservadas, pero sin presencia del arreglo clásico de cisteínas presentes en la mayoría de los péptidos de insectos (Ganz 2003) (Fig. 3).
Con base a la estructura conservada de los AMP´s reportados en Drosophila, se realizó un análisis comparativo para demostrar el arreglo espacial de las cisteínas presentes en la hebraina y en otras especies de garrapatas. Los resultados mostraron regiones de cisteína con un arreglo espacial diferente al presente en Drosophila melanogaster (D. melanogaster), pero muy similar al reportado para la Microplusina (Soneshine et al. 2013) (Fig. 4). Estos resultados demuestran la gran similitud que existe entre los AMP´s de garrapatas de otras especies, y en contraste, la diferencia que existe entra los que se han reportado para el caso de los insectos (Ganz 2003).
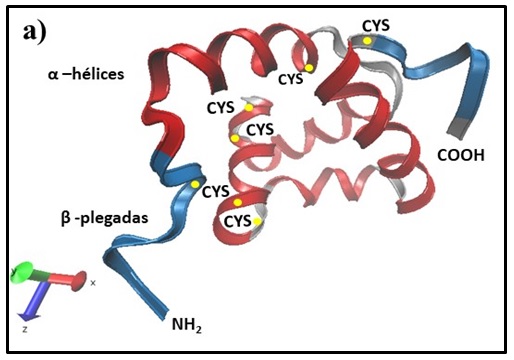 |
Predicción de la estructura en 3D de la hebraina con el programa Swiss-Model y VMD 1.9.3. El modelo muestra un arreglo estructural de las β- plegada y α- hélice, con la presencia del arreglo de 6 residuos de cisteínas conservados similar al del AMP microplusina (Aguilar-Díaz et al. 2018). |
 |
Análisis del arreglo espacial de los residuos de cisteína de la hebraina de R. microplus, comparada con la secuencia de la hebraina de D. melanogaster y la de otras especias. Se muestra el alineamiento de cinco secuencias homólogas de hebraina y la de la microplusina. Los rectángulos indican el arreglo espacial Cys1-Cys4, Cys2-Cys5 y Cys3-Cys6, idéntico al reportado para la microplusina, y similar entre las diferentes especies de garrapatas. |
2.4. Análisis filogenético de la secuencia de la hebraina de R. microplus
Para la determinación de las distancias evolutivas presentes entre las diferentes secuencias de la hebraina, se realizó un análisis de máxima verosimilitud basado en el método evolutivo de Jones-Tylor.Thornton (JTT), con la secuencia de Pseudocercospora musae (P. musae) como grupo externo (>tr|A0A139I9C2|A0A139I9C2_9PEZI), y un bootstrap de 500. Los resultados mostraron una gran similitud de la secuencia de la hebraina de R. microplus, con la de otras especies de garrapatas, presentándose como un grupo monofilético, agrupado con Rhipicephalus pulchellus (R. pulchellus), Rhipicephalus appendiculatus (R. appendiculatus), Rhipicephalus zambeziensis (R. zambeziensis) y un grupo hermano donde se agrupa Rhipicephalus sanguineus (R. sanguíneus) (Fig. 5).
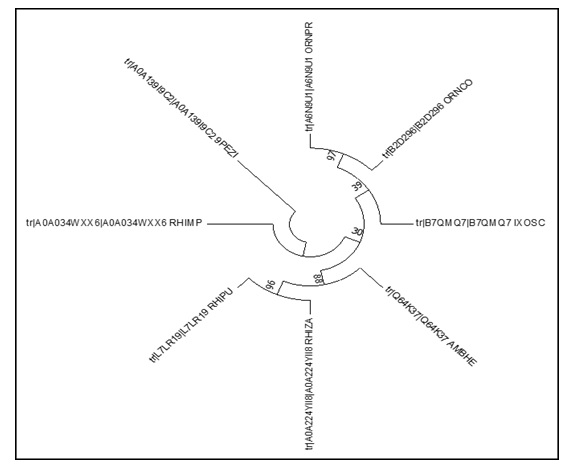 |
Análisis filogenético de la secuencia de la hebraina de R. microplus mediante un análisis de Máxima Verosimilitud y el método evolutivo de Jones-Taylor-Thornton. La imagen muestra un árbol filogenético con un bootstrap de 500. Los números cercanos a los nodos muestran los replicados del bootstrap. Como grupo externo se utilizó la secuencia de P. musae (>tr|A0A139I9C2|A0A139I9C2_9PEZI). |
3. Discusiones y conclusiones
Los resultados obtenidos en este trabajo, contribuyen a la elolucidación de los mecanismos involucrados en la respuesta inmune de R. microplus, ante el reto con una bacteria que trasmite (Anaplasma), y una patógena para la garrapata. Al respecto, se demuestra la alta plasticidad que posee la respuesta protectora, donde dos bacterias patógenas (Gram positivas y Gram negativas), producen expresión diferencial de un AMP, corroborando la idea que, dependiendo del patógeno, es el tipo de respuesta que se genera. Se muestra por primera vez, la presencia de un péptido antimicrobiano tipo hebraina presente en el genoma de R. microplus, lo cual, representa un avance el conocimiento de los AMP´s en la biología de la garrapata. Por otra parte, la estructura de la hebraina es atípica a la estructura y arreglo de los residuos de cisteínas reportados en los insectos, e interesantemente, como previamente lo reportó Soneshine en 2013, el arreglo de dichos residuos, corresponde a la estructura de la microplusina, uno de los péptidos más estudiados en R. microplus. Esto sugiere que muchos de los péptidos presentes en la garrapata, podrían mantener esta estructura conformacional. Todos estos hallazgos encontrados, ponen al descubierto el papel que tienen los AMP´s en la respuesta inmunológica de la garrapata. En este sentido, muchos autores proponen a estos péptidos como posibles agentes terapéuticos en el tratamiento de algunas enfermedades infecciosas, mostrando su alto potencial terapéutico y sinérgico en presencia de algunos antibióticos. En relación, algunos trabajos ponen en relieve el papel de los AMP´s, como inmunomoduladores de la respuesta inmune en algunas enfermedades infecciosas, donde péptidos como el LL-37, es capaz de neutralizar las endotoxinas bacterianas (LPS), inhibiendo la secreción de citocinas proinflamatorias y evitando así, la destrucción del tejido. Adicionalmente, se ha demostrado la actividad antifúngica del péptido Cm-P5 sobre cultivos de Candida albicans. Las defensinas en general, muestran actividad antiviral contra virus desnudos y los que se encuentran rodeados por una capside proteíca, aunque esta respuesta en medida por diferentes mecanismos. Esta defensa se basa en la disgregación proteica de la envoltura, o bien, uniéndose a receptores presentes en la capside evitando la unión a su célula blanco. Esta gran diversidad de mecanismos de acción, coloca a los AMP´s, como buenos candidatos de blancos terapéuticos. Por otra parte, a la fecha se sabe que parte de la respuesta inmunológica se encuentra ligada estrechamente a factores endocrinos y neurológicos, sin embrago, en el caso de las garrapatas se conoce muy poco al respecto de esto. Por lo anterior, es de considerar el papel que tienen algunas hormonas sobre el curso de la infección, y como algunos parásitos son capaces de modular esta respuesta en un proceso conocido como tras-regulacion (Escobedo et al. 2005; Aguilar-Díaz et al. 2015). En este sentido, sería interesante el considerar las infecciones de A. marginale que vienen de un hospedero macho, o de un bovino hembra. Aquí nos descartamos la idea de que la regulación de la respuesta inmune de garrapatas infectadas con sangre de bovino hembra o macho, sea diferencial. No obstante, sería otro problema que se debe abordar con mayor detenimiento. Finalmente, los análisis filogenéticos, y a pesar de compartir el mismo arreglo en los residuos de cisteínas, muestran la secuencia de la hebraina de R. microplus como un grupo monofilético, acomodado en un grupo que comparten R. pulchellus, R. appendiculatus, R. zambeziensis.
4. Futuras direcciones
Estos hallazgos en conjunto, contribuyen al avance en el conocimiento y elucidación de las interacciones vector-patógeno y en la biología de la garrapata R. microplus, con el fin de la búsqueda de nuevos blancos terapéuticos que, a futuro, contribuyan al control de esta plaga de gran impacto nacional.
Agradecimientos
Los autores agradecen al MVZ Jesús Francisco Preciado de la Torre por su asistencia para la infección y manutención de bovinos infectados con Anaplasma sp, y al Dr. Sergio Rodríguez Camarillo, por su disponibilidad para el uso de la cepa de A. marginale de la Unidad de Anaplasmosis del CENID-SAI.